Forschung
Degenerative Prozesse in biologischen Systemen, wie etwa programmierter Zelltod (z.B. Apoptose) und Altern, sind wichtige Arbeitsschwerpunkte der Entwicklungsbiologie. Das Verständnis der Grundlagen dieser komplexen Prozesse ist von großer Bedeutung und hat bedeutende Konsequenzen. So ist beim Menschen ein fortgeschrittenes Alter der wichtigste Risikofaktor zur Ausprägung verschiedener leichterer Funktionsstörungen bis hin zu schwersten Erkrankungen (z.B. Alzheimer und Parkinsonerkrankung, verschiedene Formen von Krebs). Für die Entwicklung effizienter Strategien gegen diese Erkrankungen ist das Verständnis der molekularen Grundlagen biologischen Alterns eine ganz entscheidende Voraussetzung. Solche Interventionen sollen dazu dienen, die Qualität des Lebens auch in fortgeschrittenem Alter zu verbessern. In einer „ergrauenden“ Welt mit immer größeren Zahlen älterer Menschen kommt der Alternsforschung eine immer wichtigere Funktion zu.
Da Altern auf ein sehr komplexes Netzwerk verschiedener zellulärer Stoffwechselwege und Interaktionen zurückgeht, sind in der experimentellen Alternsforschung Untersuchungen nicht ausschließlich auf den Menschen beschränkt, sondern erfolgen an einer Reihe verschiedener Modellsysteme. Modellsysteme wie Hefen, filamentöse Pilze, Fliegen, Würmer oder aber auch Zellkulturen sind von geringerer Komplexität als Säugetiere. Der Vergleich der Daten, die auf die Untersuchung dieser verschiedenen Systeme zurückgehen, bietet die größte Chance, die basalen Mechanismen degenerativer Prozesse biologischer Systeme allgemein zu erarbeiten. Die Gruppe Molekulare Entwicklungsbiologie von Herrn Prof. Osiewacz arbeitet an folgenden Schwerpunkten:
| Mitochondriale Qualitätskontrolle | |
| Oxidativer Stress, Mitochondrielle Dynamik, Proteostase, Mitophagie | |
| Rolle des programmierten Zelltods bei degenerativen Entwicklungsprozessen des Hyphenpilzes Podospora anserina | |
| Molekulare Mechanismen der Kupferhomöostase |
Förderung
Die experimentellen Arbeiten werden durch Mittel des Landes Hessen (LOEWE IPF, LOEWE Ub-Net,
DynaMem, ATG71), Sachmittel der Deutschen Forschungsgemeinschaft (Bonn, Bad-Godesberg, Deutschland ) im Rahmen von Einzelförderungsprojekten, im Sonderforschungsbereich SFB 1177 "Selektive Autophagie" und im "Cluster of Excellence - Macromolecular Complexes", durch die Europäische Union (MiMage, Proteomage, Linkage) und das BMBF (GerontoMitoSys) gefördert.
 |
 |
|

MITOCHONDRIALE QUALITÄTSKONTROLLE
Die in diesem Bereich angesiedelten Projekte verfolgen die Erforschung grundlegender Mechanismen der mitochondrialen Qualitätskontrolle. Mitochondrien dienen als „Kraftwerke der Zelle“ und sind neben der Energiebereitstellung in Form von ATP auch für die Synthese von Aminosäuren, die Herstellung von Eisen/Schwefel-Clustern und die Fettverbrennung verantwortlich. Jedoch sind Mitochondrien auch die Hauptquelle für reaktive Sauerstoffspezies (ROS), die durch eine fehlerhafte Elektronenübertragung innerhalb der Atmungskette entstehen und durch ein Netzwerk verschiedenster Reaktionen miteinander verbunden sind (Hydroxylradikale, Superoxid und Wasserstoffperoxid). In geringen Konzentrationen dienen ROS als Signalmoleküle, in höheren Konzentrationen führen sie hingegen zu Schäden der zellulären Bestandteile (DNA, Lipide und Proteine). Durch die Akkumulation der Zellschäden und die daraus resultierenden Beeinträchtigungen für die Zelle werden ROS für den altersabhängigen Funktionsverlust der zellulären Komponenten verantwortlich gemacht. Zum Schutz vor ROS existieren verschiedene Mechanismen, die nicht nur aufeinander folgend, sondern auch parallel aktiv sind.
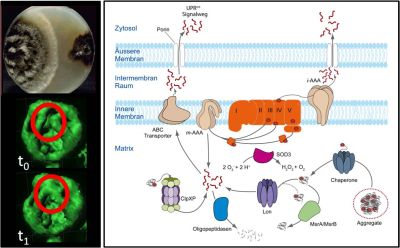
Abb. 1: Links oben ist eine wachsende Kultur von Podospora anserina (links) neben einer Kultur im seneszenten Stadium (rechts) abgebildet. Darunter sind zwei Zeitpunkte einer 4D-Mikroskopieaufnahme gezeigt. Mitochondrial lokalisiertes GFP wurde zur räumlichen und zeitlichen Beobachtung der mitochondrialen Dynamik verwendet. Die Aufnahmen entstanden an einem CLSM mit "Spinning-Disk"-Modul. Rot markiert ist ein Bereich der Mitochondrien, der von Zeitpunkt t0 zu t1 eine Fusion zeigt. Rechts sieht man einen schematischen Querschnitt durch ein Mitochondrium. Neben der Entgiftung von ROS durch SOD3 und dem Auflösen von Proteinaggregaten wird auch der Abbau von geschädigten Proteinen durch Proteasen gezeigt.
Kontakt
Prof. Dr. Heinz D. Osiewacz
Biozentrum, Campus Riedberg
Max-von-Laue-Str. 9
60438 Frankfurt am Main
F +49 69 798-29363
E osiewacz@bio.uni-frankfurt.de
Sprechzeit
nach Vereinbarung

Our website in English